Содержание
Введение 4
Глава 1. Природная обстановка позднего плейстоцена Русской равнины 8
1.1. Основные гипотезы 8
1.2. Хронология позднего плейстоцена 11
1.3. Основные черты развития природы позднего палеолита Русской равнины 12
1.3.1. Глобальное похолодание и покровное оледенение 12
1.3.2. Сочетание направленных и ритмичных изменений
природных компонентов 13
1.3.3. Высокая скорость эволюции природных компонентов 14
1.3.4. Метахтонность природных процессов 15
1.4. Особенности палеогеографических условий Восточноевропейской
равнины 16
1.4.1 Климатические условия 16
1.4.2. Эдафические условия 17
1.4.2.1. Геоморфологические особенности 18
1.4.2.2. Почвенные условия 14
1.5. Основные направления развития биоты Восточноевропейской равнины 19
1.5.1. Особенности организации вюрмских биоценозов
Русской равнины 19
1.5.2. Растительность и флора позднего вюрма 22
1.5.3. Фауна позднего вюрма Русской равнины 26
Выводы 30
Глава 2. Главные черты развития населения и хозяйства позднепалеолитических
общин Восточно-Европейской равнины 31
2.1. Палеолитические культуры центра Русской равнины 31
2.2. Особенности природопользования палеолитических общин центра
Русской равнины 34
2.2.1. Хозяйственно-культурные типы (ХКТ) палеолита Русской равнины 34
2.2.2. Особенности охоты палеолитического населения Русской равнины 37
2.2.3. Ранжирование охотничьей фауны по размерным классам 39
2.2.4. Реконструкция особенностей питания палеолитического населения
Русской равнины 40
2.3. Анализ особенностей природопользования модельных ХКТ 42
2.3.1. Особенности ХКТ кочующих охотников на стадных животных 43
2.3.2. Особенности ХКТ оседлых охотников на мамонта 44
2.4. Расчет антропогенной нагрузки на популяции пищевых доминантов для
модельных ХКТ позднего палеолита Русской равнины 45
Выводы 52
Глава 3. Некоторые особенности трансформации позднеплейстоценового
фаунистического комплекса Восточно-Европейской равнины 53
3.1. Количественное и пространственное изменение видового обилия
и размещения охотничьей фауны на позднепалеолитических поселениях
Восточно-Европейской равнины 53
3.1.1. Видовое обилие костного материала охотничьей фауны на
поселениях старше 33000 лет 53
3.1.2. Видовое обилие костного материала охотничьей фауны на
поселениях возрастом 33000 - 24000 лет назад 55
3.1.3. Видовое обилие костного материала охотничьей фауны на
поселениях возрастом 24000 -18000 лет назад 57
3.1.4. Видовое обилие костного материала охотничьей фауны на
поселениях возрастом 18000 - 12000 лет назад. 58
3.1.5. Видовое обилие костного материала охотничьей фауны на
поселениях возрастом 12000 - 9000 лет назад 59
3.2. Изменения видового обилия костного материала основных видов охотничьей
фауны на поселениях Восточно-Европейской равнины, в течение пяти
периодов позднего плейстоцена 60
3.3. Анализ изменения ареалов и обилия костного материала основных
экологических группировок “охотничьей фауны” поселений
Восточно-Европейской равнины 63
3.4. Анализ изменения соотношений животных основных трофических
уровней охотничьей фауны Восточно-Европейской равнины 69
3.5. Пространственный анализ изменения состава доминантов в охотничьей
фауне поселений Восточно-Европейской равнины в разные периоды вюрма 70
Выводы 75
Заключение 76
Список литературы 77
Приложение 91
Многие волнующие вопросы науки не могут быть решены,
если не оглянутся на вчерашний день Земли,
природы земной поверхности и обитающего на ней
человеческого общества.
К. К. Марков.
Введение
Проблема взаимоотношения природы и общества всегда была и остается основополагающей. На различных этапах развития она решалась в соответствии с совершенствованием методик и изменения взглядов на ключевые вопросы, как в археологии, так и в палеогеографии. Современное состояние проблемы зависит как от уровня развития разных наук, так и от возможностей исследователя извлечь необходимую информацию для комплексного анализа.
Исследований, посвященных трансформации позднеплейстоценовых биоценозов, немало. Однако, обобщающих работ, в которых рассматриваются изменения позднеплейстоценовых биоценозов в широком временном интервале недостаточно. В настоящее время, с развитием информационных технологий, решение проблем взаимоотношения природы и общества возможно с использованием систем управления базами данных (СУБД) и геоинформационных систем (ГИС), которые позволяют систематизировать разнообразную пространственную и временную информацию. Особенность данной работы заключается в том, что большие массивы накопленной информации по данной проблеме были инвентаризированы современными компьютерными методами.
Основным объектом исследования в работе является комплекс промысловых видов млекопитающих, обнаруженных на палеолитических поселениях восточно-Европейской равнины, получившего название “охотничья” фауна. Анализ особенностей трансформации охотничьей фауны может способствовать объективной оценке причин, инициировавших деструктивные тенденции в дальнейшем развитии позднеплейстоценовой фауны. Изучение охотничьей фауны позволяет, с одной стороны выявить особенности природопользования палеолитических общин, а с другой - не исключает возможность для экологических исследований состава и структуры фауны промысловых животных. Группировка экологических факторов и анализ характера и роли, главных из них (природных и антропогенных) в эволюции экосистем позволит прогнозировать специфику развития территории в будущем.
Цель работы - охарактеризовать вклад природных и антропогенных факторов в трансформацию позднеплейстоценовых экосистем Восточно-Европейской равнины.
Задачи
- Оценка возможного влияния хозяйственной деятельности на состояние популяций основных объектов охоты.
- Создание базы данных основных видов охотничьей фауны с палеолитических поселениях Восточно-Европейской равнины и серии ретроспективных карт, отражающих качественные и количественные изменения её состава.
- Анализ изменения структуры и ареалов видов охотничьей фауны позднеплейстоценовых поселений Восточно-Европейской равнины.
- Выявление роли природных и антропогенных факторов в трансформации позднеплейстоценовых экосистем Восточно-Европейской равнины.
Методы
Основной проблемой при проведении комплексного анализа изменения распространения видов животных в прошлые эпохи является разрозненность фактического материала. Данные о находках костей животных рассеяны по археологическим, палеонтологическим, геологическим и палеогеографическим работам. Сводные работы редки и труднодоступны. Автором составлена База Данных (БД) находок остатков 52 видов млекопитающих, обнаруженных на палеолитических поселениях и относимых к охотничьей фауне. Датировки находок, составляющих БД, попадают в интервал 100000 - 10000 лет назад. Материалы, представленные в работе, охватывают территорию Восточной Европы. БД была организована в системе Access и состоит из 4 таблиц:
“Geopoint” - таблица включает информацию о расположении 403 местонахождений позднеплейстоценовой фауны (область, район, ближайший населенный пункт, координаты).
“Age”- в таблице собранны радиоуглеродные (около 250), археологические и геологические датировки местонахождений позднеплейстоценовой фауны
“Fauna” - таблица включает сведения об обнаружении 52 видов млекопитающих на позднепалеолитических поселениях Восточной Европы. Более чем для половины местонахождений приведены количественные оценки минимального числа видов или числа костных остатков
“Referens” - в таблице представлены печатные источники, использовавшиеся для составления БД.
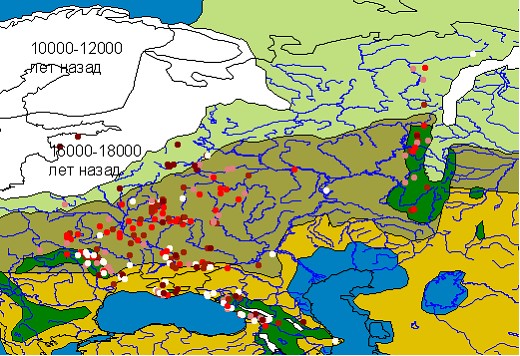
Данные, собранные в БД, были визуализированы в виде геоинформационной системы в среде ArcView. Анализ особенностей изменения структуры охотничьей фауны проводился для пяти периодов позднего плейстоцена, выделение которых соответствует периодизации освоения Русской равнины человеком (Величко, Куренкова, 1994) и палеогеографическим схемам климатических пульсаций вюрма (Динамика.., 2002). Материал ранжирован по следующей схеме: средний вюрм (мустье, 100000-33000 лет назад), брянский интервал (ориньяк, 33000-24000), верхневюрмский пессимум (солютре, 24000-18000), позднеледниковье (мадлен, 18000-12000), послеледниковье (переход к мезолиту, 12000-9000). На рис. 1.1 (приложение) представленно территориальное размещение местонахождений охотничьей фауны на территории Русской равнины.
В работе проведена оценка обилия видов, обнаруженных в составе охотничьей фауны позднепалеолитических поселений. В связи с отсутствием единой схемы оценки видового богатства обилия по костному материалу, в работе проведен синтез известных в настоящее время способов определения видового обилия по костному материалу. В отечественной палеонтологии принято несколько способов оценки видового богатства ископаемой фауны:
- Определение минимального числа особей каждого вида, исходя из обнаруженного костного материала.
- Подсчет всех костных остатков, которые можно определить до вида либо другой систематической группы.
- Словесная оценка обилия.
Словесная оценка обилия костного материала на позднепалеолитических поселениях Русской равнины использовались ранее в некоторых работах (Громов, 1966; Иванов, 1980). В ряде работ, особенно ранних, оценка обилия костного материала не производилась. Отмечалось только присутствие или отсутствие вида.
Автором проведена оценка видового обилия костного материала по 5балльной шкале. Каждому баллу соответствует количественный или словесный критерий оценки видового обилия костного материала по разным схемам. В данной работе, в случаях, когда для обнаруженного вида отсутствует информация, позволяющая предположить его обилие, виду присваивался минимальный коэффициент обилия, принятый в работе, - единица.
Таблица 1.1.
Сопоставление различных схем оценки видового разнообразия по ископаемому костному материалу
Балл
|
Минимальное число особей вида
|
Число костных остатков одного вида
|
Словесная оценка вида в костном
материале.
|
1
|
1
|
1 - 10
|
Отмечен
|
2
|
2 - 4
|
11 - 30
|
Много
|
3
|
5 - 8
|
31 - 60
|
Очень много
|
4
|
9 - 12
|
61 -100
|
-
|
5
|
Более 13
|
Более 100
|
-
|
Методы исследования
1. Анализ литературных и электронных источников
2. Создание БД
3. Создание электронных карт
4. Статистические методы обработки БД
Глава 1. Природная обстановка позднего плейстоцена Русской равнины
1.1. Основные гипотезы
Ключевым событием плейстоцена стала смена температурного тренда на рубеже плиоцена и плейстоцена (Брукс, 1956; Четвертичный период, 1968а; Дашкевич, 1969; Котляков и др., 1984; Зубаков, 1986; Лазуков, 1989). Наметившееся вслед за этим глобальное похолодание изменило природную обстановку во многих регионах мира и особенно ярко проявилось в умеренных широтах (Брукс, 1956; Четвертичный период, 1968а; Котляков, 1979; Тарлинг, 1980; Сукс, 1986). Описанное событие в настоящий момент признано большинством исследователей, однако, в степени оценки отклика географической оболочки на него существуют серьезные разногласия.
В настоящее время наиболее полно разработаны две основные гипотезы развития биосферы в плейстоцене: антигляциальная (Пидопличко, 1954; Линдберг, 1955; Калякин, 2001) и гляциальная (Герасимов, Марков, 1939; Четвертичный период, 1968а, Величко, 1994; Динамика …,2002).
Антигляциальная гипотеза представляет собой группу представлений (делювиальные, дрифтовые, элювиальные), которые отвергают возможность формирования покровного оледенения в плейстоцене на территории северных материков. По опубликованным данным, свое начало антигляциальные гипотезы ведут с работы Аренсвальда вышедшей в 1775 году, в которой он объяснял занос эрратических валунов северной Европы водами всемирного потопа (Герасимов, 1939). В начале 19 века Плейфер оформил группу подобных взглядов в элювиальную гипотезу происхождения аккумулятивных плейстоценовых образований Европы (Меннер и др.,1986). В дальнейшем эти взгляды послужили основой для дрифтовой гипотезы Ч. Лейеля (ледниково-морсково происхождения эрратических валунов), ставшей главным ядром антигляциальной гипотезы (Герасимов, Марков, 1939; Бердников, 1983; Меннер и др., 1986; Неймаер, 1994). Российская антигляциальная школа ведет свое начало с работ П. С. Палласа, объяснявшего в 1778 году наличие сибирских валунов и костей мамонта действием гигантского наводнения (Меннер и др., 1986; Коэн, 1996). В 19 веке его идеи были поддержаны Ф. Н. Чернышовым, А. А. Штукенбергом, И. С. Поляковым, сформулировавшими отечественную гипотезу “о морских валунных суглинках на севере России” (Меннер и др., 1986). В 20 веке доводы сторонников антигляциальной гипотезы получили фактическое подтверждение в работах некоторых геоморфологов (Чувардинский, 1967; Цыганов, 1972; Цуриков, 1972; Боландин, 1986), палеоботаников (Вульф, 1933; Кожаринов, Морозова, 2001),и палеозоологов (Шарфф, 1889; Пидопличко, 1951, 1954; Линдберг, 1955; Калякин, Турубанова, 2002).
В современное время основные положения антигляциальной гипотезы выглядят следующим образом:
- В течение всего плейстоцена значительные покровные оледенения на территории северных материков отсутствовали.
- Главные события плейстоцена, это периодические средне и высокоамплитудные (100-300 метров) колебания земной коры, сопровождавшиеся крупными трансгрессиями и регрессиями, значительно менявших природную обстановку северных материков.
- Климат Голарктики в плейстоцене эволюционировал от тропического и субтропического к бореальному (близок современному), а его изменение не имело характер глобального катаклизма.
- Главное ядро биоты северных материков имеет автохтонное происхождение и берет свое начало в третичных биоценозах, а миграционных компонент имеет второстепенное значение.
- Эволюция биоты Голарктики носила направленный характер и представляла собой смену теплолюбивых видов, главным образом, бореальными. Роль арктических видов на протяжении всего плейстоцена не была главной.
Гляциальная гипотеза ведет свое начало с середины 18 века, когда Соссюр опубликовал работу, посвященную древнему оледенению Альп (Герасимов, 1939). В 19 веке эти взгляды, развитые в работах классиков гляциализма Агассица, Ф. Б. Шмидта, П. А. Кропоткина, Ч. Д. Черского, О. Торреля, делают гипотезу господствующей схемой развития биосферы в плейстоцене (Герасимов, 1939; Бердников, 1983; Меннер и др., 1986; Шанцер, Вангэнгейм, 1986).
В гляциальной гипотезе выделяется три основных направления. Исторически первым в их ряду был моногляциализм. Его представители отстаивают существование одного устойчивого похолодания в плейстоцене, завершившегося однократным формированием покровного оледенения северных материков (Колесник, 1939; Серебрянный, 1980). Популярное в настоящее время направление - полигляциализм - основано на схеме периодизации Пенка - Брюкнера о трехкратном оледенении Альп, чередовавшихся с теплыми и влажными межледниковьями (Четвертичный период, 1968а). В дальнейшем схема была расширена до 4-5 гляциалов (Рослых, 1986; Лазуков, 1989). Активное развитие полигляциализма в 20 веке привело к появлению его крайнего варианта плюрогляциализма (по словарю Л. И. Муруашмили, 1980). Н. С. Болиховска (1995) выделяет в плейстоцене Восточной Европы 8 ледниковий и 9 межледниковий. В этом случае повышается ранг стадиалов и интерстадиалов до гляциалов (Муруашвили, 1980).
Основные положения гляциальной гипотезы можно представить следующими положениями:
- В течение плейстоцена происходила периодическая смена периодов глобального похолодания и потепления климата, находивших прямое отражение в изменении природной обстановки Земли.
- Следствием глобальных климатических пульсаций стало чередование гляциальных эпох с интергляциальными.
- Ключевым событием гляциальных эпох явилась активизация горных ледников и формирование равнинных оледенений умеренных широт обоих полушарий.
- Изменение климата плейстоцена имело ритмичный характер и развивалось как серия направленных пульсаций, осложненных флуктуациями низших рангов, менявших климатические условия от бореальных и субтропических (интергляциалы) к арктическим и субарктическим (гляциалы).
- Основное направление развитие биоты плейстоцена - формирование группы холодолюбивых видов за счет перегруппировки или вымирания основного ядра автохтонных видов и включения в ценозы адвективных видов.
- В плейстоцене происходили многократные миграции целых биоценозов в южном направлении в гляциальные эпохи и в северном - в интергляциальные.
Природные следствия ледниковой гипотезы получили неоднозначное толкование в работах её сторонников. Многие исследователи определяют природные условия гляциальных эпох как “суровые” или ”экстремальные” (Герасимов, Марков, 1939; Брукс, 1952; Величко, 1973; Ушаков, Ясманов, 1984; Величко и др., 1999). По мнению других роль покровного оледенения была локальной и его влияние на природную обстановку преувеличено. Еще в 1889 году Шарфф указывал на противоречие имеющегося палеонтологического материала с реконструкциями экстремальных природных условий плейстоцена. В 1933 году Вульф указывал на существование теплолюбивой флоры на территории Европы в ледниковья плейстоцена. М. А. Мензбир, опираясь на палеонтологические данные, полагал, что достоверно можно говорить только о том, что условия гляциальных эпох не были тропическими (цит. по: Четвертичный период, 1968в). И. Г. Пидопличко (1954) отмечал: ”Это охлаждение климата, вполне доказываемое палеонтологически, не было слишком губительным, однако оно обусловило появление в северной и средней полосе Европы условий, способствовавших проникновению на юг ряда северных видов”. П. И. Дорофеев делает схожий вывод: “Миндельские и вюрмские флоры всегда комплексные и даже смешанные. Только бореально - арктические (а иногда только таежные) элементы этих флор могут толковаться как доказательства ледниковой эпохи, причем не самого оледенения, а только несколько иного климата по сравнению с современным” (цит. по: Пидопличко, 1967). Л. Р. Серебрянный (1980) так же высказывает сомнения в грандиозности природных изменений вюрма. Он обращает внимание на то, что по количественному перераспределению соотношения “северных” и “южных” видов, которые периодически происходили в вюрме, нельзя делать выводы о смене межледниковый и ледниковий.
1.2. Хронология позднего плейстоцена
Необходимо отметить, что почти все современные палеогеографические (Величко, 1961, 1973, 1994; Гричук, 1989; Лазуков, 1989; Динамика.., 2002) и археологические (Природа и древний.., 1981; Рогачев, Аникович, 1984; Величко, Куренкова; 1994) схемы развития природы и общества в плейстоцене выполнены на основе гляциальной гипотезы. В основе геохронологических шкал также лежат представления о чередовании термоэр и криоэр (Рослых, 1986; Динамика.., 2002). По ним поздний плейстоцен включает в себя вюрмский (валдайский) мегакриохрон (117 - 15 тысяч лет назад). В словаре Л. И. Маруашвили (1984) нижняя границы вюрма определена в 185- 70 тыс. лет назад, а верхняя граница в 11-10 тыс. лет назад. Н. А. Хотинский (1981) определяет рубеж плейстоцена и голоцена в 10300 лет, а М. И. Нейштадт (1957) в 12000 лет назад. В. А. Зубаков (1986) рассматривает вюрм как серию ортоклиматем (гляциалов и интергляциалов). Е. А Спиридонова (1991) называет валдайский ледниковый этап сложным климатическим ритмом, осложненным климатическими колебаниями разных рангов. Вопрос о статусе рангов до настоящего времени однозначно не решен. В таблице 1.2. представлены некоторые схемы периодизации позднего вюрма, используемые в настоящее время.
Таблица 1.2.1.
Сопоставление разных схем периодизации позднего вюрма
Возраст тыс. лет.
назад
|
Почвенно -
стратиграфическая
|
Археологическая
|
Палеогеографические
|
Советская
|
Цен. Европа
|
Зап. Европа Приднестровье
|
Ландшафтно-климатические.., 1999
|
Палеолит СССР,
1984
|
Лазуков, 1989
|
Муруашвили,
1984
|
Флон,1980,
Болиховская,
1995
|
Маркова, 2002
|
10
25
50
75
|
Супоневский лес
Алтыновский лес
Трубчевский почвенный уровень
Деснинский лес
Брянская почва
Хотылевский лес
Смоленский криогенный горизонт
Мезинский
почвен. комплекс
|
Азиль
Мадлен
Солютре
Ориньяк
Периге, селет
Финальное
Мустье
Мустье
|
осташковский
гляциал
молого-шекснинское
интергляциал
калининский
гляциал
|
Даун
Гигниц
Бюль
|
Мол. Дриас
Аллерёд
Др. Дриас
Бёллинг
Ласко
Штилфрид В
Арсу
Денекамп
Хенголо
Поперинге
Оддераде
Брёруп
Амерсфоорт
|
Днестровский
Молодовский
Байловский
Кишлянский
Кетросский
|
1.3.3. Высокая скорость эволюции природных компонентов
Многие исследователи обращают особое внимание на высокую скорость эволюции природной среды в некоторые интервалы вюрма (Хотинский, 1981; Украинцева, 1991; Величко, 1973,1994). Подобные явления определяются как ”палеогеографические рубежи, в течение которых происходят качественные изменения, имеющие характер скачков” (Долуханов, Пашкевич, 1977). Переходы поздневюрмских природных систем из одного состояния в другое проходили часто стремительно до финального палеолита без ущерба для ее биотических компонентов. Переход от бореального климата через субарктический к арктическому произошел за 3000 лет (Природа и человек, 1984). По данным Флона (1980), пульсации среднегодовых температур в 5-6 градуса могли происходить за столетие. Особенно ярко ускорение скорости эволюции природных процессов проявилась во второй половине вюрма. В Западной Европе за период с 16000 до 10000 лет назад рост температур самого теплого месяца составил 18 градусов (Природа и человек..,1984). М. Г. Гросвальд (1989) указывает на рост среднегодовых температур на рубеже голоцена в Гренландии на 20 є С. По данным палеонтологов отступающий ледник прошел путь от центра Валдайской возвышенности до Скандинавии за 2-4 тыс. лет (Морозова, Кожаринов, 2001). Период окончательного распада покровного ледника составил 1500 лет (Зубаков, 1986).
Для объяснения столь быстрого протекания природных процессов М. Г. Гросвальд (1999) предложил гипотезу гравитационного коллапса ледника, по которой рост и распад ледников мог осуществляться в геологические ”мгновения” (сотни лет). По данным палеонтологов, для развития биоты вюрма можно выделить два рубежа природных преобразований, имеющих черты ускорения эволюции природных компонентов: 40000 и 13000-8000 лет назад (Верещагин, 1974; Дмитров, Белокобыльский, 1989; Украинцева, 1991) .
1.3.4 Метахтонность природных процессов
Развитие природного процесса позднего плейстоцена Евразии называют метахтонным (Долуханов, Пашкевич, 1977; Хотинский, 1981; Величко, 1994). В пределах Голарктики часто крупнейшие природные события не совпадали во времени. Некоторые исследователи отмечают изменение характера действия экологических факторов в разные периоды вюрма. Так же необходимо отметить, что период роста покровного ледника составлял 10 тыс. лет, а деградация произошла стремительно - за 4 тыс. лет (Зубаков, 1986; Хотинский, 1981). Максимальное продвижение льдов в северном полушарии отмечалось не в стадиалы, а в интерстадиалы (32 и 14 тыс. лет назад) (Флон, 1980). Он же указывает на наличие в вюрме 5 похолоданий примерно сравнимых по масштабам, при этом образование покровного оледенения отмечалось только в одном из них. Отмечается запаздывание проявления природных событий в Восточной Европе по сравнению с западом материка. Максимум похолодания в западном секторе Европы отмечен в интервале 28000-26000 лет назад, а в восточном - на 10000 лет позже (Природа и человек, 1984). Природные преобразования в Западной Европе начались раньше и протекали интенсивнее, чем в Восточной (Флон, 1980; Серебрянный, 1980; Хотинский, 1981). Палеонтологи отмечают (Соффер, 1993), что трансформация мамонтового фаунистического комплекса в западной Европе опережало аналогичный процесс в Восточной на несколько тысячелетий. Разные секторы восточной Европы имели индивидуальные особенности развития.
В позднем вюрме западные и восточные секторы Восточноевропейской равнины, вероятно, отличались по продуктивности. Установлено, что процент содержания гумуса в Ароповичах (Украина) 1,18%, на 400 км. восточнее на Калачской возвышенности (Ростовская область) - 0,5%. Мощность почв также уменьшалась с запада на восток и с юга на север (Лессово-почвененные.., 1997).
1.5.2. Растительность и флора позднего вюрма
Обзор палеогеографической литературы показал, что перигляциальные типы растительности Русской равнины часто реконструируются как перигляциальные степи, лесостепи, тундро-степи и тундро-лесостепи (Марков и др., 1964; Гричук, 1989; Болиховская и др., 1999). Иногда эти типы объединяются в одну группу, получившую множество названий: лессовая степь (Юрцев, 1981), криогенная савана (Верещагин, 1974), болото-степь (В.А.Ривердато, по Четвертичный период, 1965в), перигляциальная степь (Гричук, 1989), холодная степь (Н.М. Крашенинников, по Четвертичный период, 1965в), мамонтовая криостепь (Зубаков, 1986) и т.д. Б. А. Юрцев (1981) дает следующее определение такому типу растительности: “группа сообществ с содоминированием в них микротермных ксерофитов или гемикриоксерофитов, с одной стороны, криоксерофитов - с другой ”. Тундро-степь отличалась тем, что травяной ярус ее формировали в основном не мхи (как в современной тундре), а злаки; здесь складывался крайне криофильный вариант травяного биома с его высокой биомассой пастбищных копытных и хищников (Украинцева, 1993). В настоящее время установлено, что диапазон типов перигляциальных сообществ был значительно шире (таблица 1.5.1) (Болиховская, 1995; Болиховская.и др.,1999).
Таблица 1. 5.1.
Развитие растительности центра Русской равнины в поздем вюрме
Источник
|
Возраст
(тыс. лет назад)
|
Фаза развития
растительности
|
Доминирующие сообщества
|
Гричук, 1989
|
40 -25
(переходная стадия)
25-10 (криоксеротическая
стадия)
|
Перигляциальная лесостепь
Комплексная криоксеротическая растительность.
Безлесная криоксеротическая растительность
|
Луговые степи с елово-березовыми и сосновыми лесами и солонцеватые
луга с Atriplex tatarica.
Сочетание степных группировок (с Ephedra clistachya, Eurotia ceratoides) и тундровых кустарниковых сообществ (с Betula nana, Lycopodium alpinum,Selaginella selaginoides).
Господство степных ассоциаций с (Artemisia laciniata, Kochia prostrata), с присутствием солончаковых ассоциаций (Solicornia herbacea, Solsola toliosa) и компонентов тундровой растительности (Echinopsilon)
|
Болиховская,
1995
|
40-25
(брянская почва)
25-10
(лес 2)
|
Экстрогляциальная лесостепь
(парковые леса)
Перигляциальная лесостепь
|
Сосново-березовые (с липой, вязом, грабом) и березово-кедрово-елово-сосновые группировки (с вязом и дубом) и степное разнотравье.
Сосновое редколесье со степными и луговыми криоаридными и ерниковыми сообществами и большой площадью эродированных участков.
|
Типизация палиноспектров..,
1999
|
40-25
брянский интервал
25-10
(верхневалдайский пессимум)
|
Перигляциальная
Лесостепь
Перигляциальная тундра и
лесотундра
|
Елово-сосновые, сосновые, березовые, разряженные леса, чередующиеся с ерниками, разнотравно-злаковыми и осоковыми группировками.
Разряженные леса (березовые, сосново-березовые с примесью ели, лиственницы) и ерниковые заросли.
|
По видовому составу спорово-пыльцевые спектры разных районов Русской равнины часто имеют сходный плюрозональный состав. Отличия заключаются, главным образом, в наборе доминантов видов и ассоциаций.
Основные отличия перигляциальных палиноспектров лессово-почвенных формаций от современных:
Более низкое содержание термофильных элементов дендрофлоры и микрофоссилий трав (Сукачев, 1932; Гричук, 1989; Болиховская и др., 1999).
Автохтонным совмещением микроостатков тундровой, бореально-лесной и пустынно-степной флоры (Дашкевич, 1964; Серебрянный, 1980; Новенко, 2000).
Высокая доля пионерной травяной растительности (Гричук, 1989; Новенко, 2000).
Расширение ареала арктических и гипоарктических, степных и пустынных видов на 1000-1200 км. к югу и северу соответственно (Рослых 1986; Болиховская, 1995).
Заметное преобладание пыльцы трав и спор над пыльцой древесно-кустарниковых видов (Асеев, 1956; Болиховская и др., 1999).
Даже в самые холодные интервалы вюрма не подтверждается полная деградация лесного пояса. Еще Ф. Н. Мильков (1952) писал: ”Вместе с тем у нас нет никаких оснований говорить об исключительной суровости климата ледниковых эпох. Наоборот, есть все основания к тому, что бы предполагать существование на внеледниковых возвышенностях юга Русской равнины не только хвойных и мелколиственных лесов, как на низменных равнинах, но и широколиственных лесов…Не исключена возможность произрастания вблизи валдайского ледника даже широколиственных лесов”.
Л. Р. Серебрянный (1980) отмечает: “Во время последнего оледенения происходила лишь частичная перестройка растительного покрова с выживанием лесов в благоприятных условиях”. Лесная область Русской равнины в позднем вюрме представляла собой редуцированные сообщества разреженных “парковых” лесов (Авенариус и др., 1978). Амфибореальное распространение вюрмских лесов Восточно-европейской равнины с участием широколиственных видов подтверждено во многих работах (Серебрянный, 1980; Удра, 1988; Markova and all, 2000; Кожаринов, Морозова, 2001). Существуют основания полагать, что на протяжении всего вюрма в центре Русской равнины произрастали широколиственные породы (липа, вяз, возможно, дуб). Палеоботанические свидетельства этого приведены во многих работах (Асеев, 1954; Артюшенко, 1959; Губонина, 1977; Левковская, 1977; Величко и др., 1981; Удра, 1988; Невенко, 2000; Кожаринов, Морозова, 2001). В эпоху “максимального холода” широколиственные породы (вяз, дуб, липа, клен) продолжали встречаться по поймам до 49-51°сш. (Асеев, 1954; Удра, 1988). Л. П. Александрова и С. Н. Цейтлин (1965) по фауне грызунов реконструируют в поймах позднего плейстоцена центра Русской равнины заболоченные хвойно-широколиственные леса.
Нередко пыльцу термофильных листопадных пород обнаруживают на расстоянии около 100 км от предполагаемой границы ледника (Авенариус и др., 1978). Термофильная флора, отмечаемая на Русской равнине в разные интервалы вюрма, включающая кроме широколиственных пород и термофильные травы и споровые, получила название аномальная флора (Болиховская, 1995). В. П. Гричук (1989) в верхнеплейстоценовых отложениях средней части Русской равнины указывает на нахождение “показательных видов”: диффузно-рассеянные термофильные (Brasenia purpurea Mich., Aldrovanda vesiculosa L., Trapa natans L.), американские (Dullchium arundinaceum Brit.), восточно-азиатские (Osmunda cinnamomea L.), европейские (Potamogeton polygonifolius Pour., Stratiotes aloides L., Carpinus betulus L., Tilia platiphyllus Scop). А. Т. Артюшенко (1959) сообщает об обнаружении растений, относящихся к межледниковым реликтам: хвоща гигантского (Equisetum maximum), башеницы голой (Turitis glabra), лунника оживающего (Lunaria radiviva), вязеля стройного (Qoronilla etegans), пыльцеголовника белого (Cephalanthera alba). А. А, Величко и др. (1997) сообщают о нахождении пыльцы болотного теплолюбивого хмеля (Humulus lupus), в настоящее время произрастающего на Кавказе.
Вопрос о причинах успешного существования теплолюбивой флоры в холодные периоды вюрма еще не решен. Гипотеза о существовании рефугиумов теплолюбивой флоры на территории Восточноевропейской равнины высказывалась неоднократно (Артюшенко, 1959; Четвертичный период, 1965в; Марков и др., 1964; Гричук 1989; Marcova and all, 2000). В настоящее время их количество только на Русской равнине оценивается в 3 десятка (Кожаринов, Морозова, 2001). Так же обращает внимание широкое распространение на Русской равнине видов, в настоящее время ареал которых захватывает только ее окраины: Larix sibirica, Picia sibirica, Abies (Гричук, 1989).
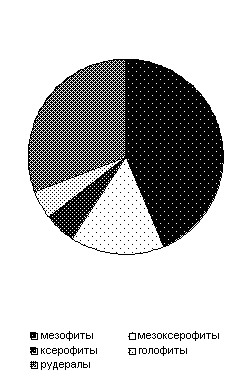
В настоящее время установлено, что перигляциальная ископаемая флора включала не менее 90 таксонов. Из них 74% бореальные таксоны (приложение, рис. 1.5.1.) (Новенко, 2000).
Эколого-ценотические исследования указывают на доминирование в перигляциальных фитоценозах двух групп: мезофитов и рудералов. Высокая доля пыльцы растений последней группы может свидетельствовать о широком распространении нарушенных субстратов. Эколого-ценотические исследования подтверждают гипотезу о высокой комплексности ландшафтов.
1.5.3. Фауна позднего вюрма Русской равнины
Развитие фауны Русской равнины имело автохтонный характер. Плейстоцен был в основном периодом формирования ареалов и экологических группировок уже сложившихся видов (Дашкевич, 1964). И. Г. Пидопличко (1956) отмечает: “фауна плейстоцена продолжает фауны гомиоцена, она ниоткуда не пришла и никуда не ушла…”. К.К.Марков и др., (1965) представляют развитие фауны плейстоцена следующим образом: “”старые” и “новые” виды, первоначально существовавшие одновременно, занимали соответствующие ландшафты или районы. Затем, в эпохи смен одного фаунистического комплекса другим, происходило перераспределение количественных соотношений между видами…”.
Эволюция в специфических условиях, привела к формированию особого эколого-фаунистического комплекса перигляциальных зон с особыми морфо-анатомическими и экологическими особенностями. Верхнепалеолитический фаунистический комплекс, свойственный позднему вюрму Русской равнины (50 - 10 тыс. лет назад), сформировался из хазарского комплекса (Алексеева, 1990). В настоящее время два этих комплекса объединяются в мамонтовый териокомплекс, ранний и поздний соответственно (Динамика.., 2002). Его составляли как панпалеоарктические, так и панголарктические виды. На Восточноевропейской равнине выделяют несколько его локальных вариантов: арктический, бореальный, северокавказский подкомплексы. Выделение подкомплексов основано, прежде всего, на изменении обилия доминантов (Давид, 1980; Динамика.., 2002).
Дифференциация мамонтового комплекса возрастала с севера на юг. С продвижением к югу возрастала доля “южных” видов, при этом “северные” всегда присутствовали (Верещагин 1979; Агаджанян, Еребаева, 1983; Динамика.., 2002). Севернее 50 є с. ш. преобладали мамонт, шерстистый носорог, северный олень, арктические грызуны. Южнее, на ряду с выше перечисленными, возрастает количество находок овцебыков, антилоп, бизона, благородных оленей и др. (Серебрянный, 1980).
Развитие позднепалеолитического фаунистического комплекса часто представляется как направленный, прогрессивный процесс, завершившийся в позднем вюрме формированием “комплекса видов глубоко адоптированных к условиям суровых биотопов плейстоцена” (Алексеева, 1990). Однако, в настоящее время можно утверждать, что эволюция фауны не была столь однозначной. Н. К. Верещагин (1974, 1979) отмечает сложный характер эволюции фауны позднего плейстоцена. В течение этого периода анатомо-морфологическое строение и, возможно, экология многих видов изменятся (Пидопличко, 1951; Верещагин, 1953, 1974, 1979, 1980; Алексеева, 1990; Украинцева 1991; Саблин, 2000). В ходе второй половины плейстоцена происходила последовательная смена нескольких адоптивных форм животных одного вида (анатомо-морфологические и эколого-физиологические изменения). Последние изменения, начавшиеся с 25-20 тыс. лет назад, закончились в позднем голоцене образованием современных форм (Алексеева, 1990; Саблин, 1997). При этом изменения, вызванные вырождением видов, образуют отдельную группу, и являются показателем вымирания вида (Верещагин, 1972; Украинцева, 1991; Алексеева1990; Сулержицкий, 1995). Собранный материал позволяет утверждать, что эти изменения часто были синхронны климатическим пульсациям и могли происходить в сравнительно короткие сроки. В. В. Украинцева (1991) отмечает: “Для плейстоценовых животных высокие скорости эволюции диктовались высокой скоростью эволюции природной среды. А так как при этом скорость смены холодных и теплых ритмов нарастала, а продолжительность их сокращалась, то качество и темпы адаптации у животных нарушались и не поспевали за эволюцией природной среды”. Высокая скорость морфологических изменений в позднем вюрме подтверждена и на мелких млекопитающих (Агаджанян, Ербаева, 1983). Выделено несколько периодов позднего вюрма, в которые активизировались адаптивные процессы в зооценозах: 40-30 и 15-10 тыс. лет назад (Николаев и др., 2000; Дмитров, Белокобыльский, 1989).
Подтверждает это и распределение радиоуглеродных датировок остеологических материалов по мамонту для северной Евразии (приложение, рис. 1.5.2).
Наибольшее число находок костей мамонтов приурочено к термоплювиальным фазам плейстоцена (40-45, 30-25 тыс. лет назад). Судя по распределению радиоуглеродных дат, численность мамонтов в периоды вюрма сильно колебалась. Резкий всплеск находок в начале голоцена, можно трактовать как свидетельство интенсивного вымирания мамонтов. Многие исследователи указывают для этих периодов на повышенную концентрацию аномалий строения костей скелета и зубов у некоторых видов животных (Верещагин, 1972; Кузьмина, Праслов, 1992; Agenbroad, 1994-1998). Измельчанию в эти периоды подверглись: мамонт, северный олень, овцебык, песец, волк, бурый и пещерный медведь и некоторые другие виды (Громов, 1966; Кузьмина, Саблин 1992; Мащенко, 1992; Саблин, 1997).
Наиболее четко эта закономерность прослеживается в изменение высоты тела половозрелых особей Mammuthus primigenius в позднем палеолите (приложение, рис. 1.5.3).
На протяжении позднего палеолита высота тела половозрелых особей мамонтов сокращается. Красной линией отмечена высота типичного позднего мамонта. Видно, что до начала брянского интервала 40-45 тыс. лет назад (потепление) высоты мамонтов превышают красную линию, а после постепенно снижается и опускается ниже неё. Возможно, это является следствием изменившейся природной обстановки.
Еще одной особенностью определенных выше периодов стало сокращение ареала для некоторых видов. В литературе приводятся сведения об уменьшении ареала мамонта, после 50 тыс. лет назад (Алексеева, 1990; Baryshnikov, 1994-1998; Динамика.., 2002), пещерного льва и гиены, сайги, пещерного медведя (Верещагин, 1974), овцебыка (Тихомиров, 1985). Во второй половине вюрма ареалы некоторых видов продолжают деградировать и популяции распадаются на локальные варианты (Алексеева, 1990; Украинцева, 1991). О. Ф. Соффер (1993) указывает, что описанный процесс начался в Западной Европе, откуда распространился на восток. По мнению археологов в этом же направлении шло расселение человека в позднем вюрме (Праслов, 1984).
В литературе отмечено для поздневюрмской фауны, нарушение возростно-половой структуры популяций, сокращение онтогенеза (Мащенко, 1992). Следствием описанных выше черт деградации популяций некоторых видов животных, могла стать возросшая смертность от естественных причин (Верещагин, 1979; Алексеева, 1990; Соффер, 1993; Исследования.., 1995), что способствовало сокращению систематического разнообразия в разных таксонах (приложение, рис. 1.5.4.).
Кроме анатомо-морфологических изменений, многие виды в течении вюрма существенно меняли свою экологию. А.Я. Тугаринов (цит. по Четвертичный период, 1965) отмечал: “ было время, когда песец был таким же арктиком, как сайга, а северный олень столь же степным, как и лошадь”. И. Г. Пидопличко (1954) указывал, что многие виды первые этапы своего развития проходили в субтропических условиях, а в настоящее время ареал их обитания захватывает тундру и северную тайгу. Н. К. Верещагин (1974, 1979), рассматривая вопрос о широких ареалах некоторых видов, называет их космополитами. По его мнению, космополитами, кроме вымерших, были и существующие в настоящее время степные, тундровые виды. Палеонтологи делают выводы об изменении особенностей питания видов на основе изучения анатомо-морфологического строения. Некоторые из них полагают, что современные тундровые виды (северный олень, овцебык), входившие в состав мамонтовой фауны, в вюрме имели строение зубной системы, указывающее на доминирование в пищевом рационе жестких (степные злаки) трав (Кузьмина, Саблин, 1992; Тихомиров, 1985). М. В. Саблин (1997) считает, что анатомические изменения, закончившиеся формирование современных видов вюрмской фауны начались не раньше 15 -17 тыс. л. н.: именно к этому времени относится появление форм близких современным. Аналогичные выводы были сделаны при изучении фауны Костенковско-Борщевского района (Верещагин, Кузьмина, 1982). Доказательством возможных изменений экологии видов, могут служить сведения о былом распространении некоторых из них (Кириков, 1966). Современные данные показывают, что особенности питания и поведения видов может меняться стремительно. За последние 200-300 лет в ряде районов Сибири пищевая специализация северного оленя менялась трижды: до одомашнивания основу его пищевого рациона составляла злаково-осоково-разнотравная растительность, затем произошел переход на “ягелеядность”, а следствием истощения пастбищ стал возврат к исходной пищевой специализации (Егоров, 1965; Сыроечковский, 1986). Подтверждением выше приведенных выводов могут стать данные по успешной акклиматизации некоторых видов, как считается, в несвойственных условиях. Интродукция северного оленя на туссоковые злаковники островов Субантарктики дала рекордную плотность: 61 особь/кмІ (Сыроечковский, 1986).
Выводы
- Развитие позднеплейстоценовых биоценозов Восточноевропейской равнины происходили на фоне периодических климатических пульсаций. Переход из одного состояния в принципиально иное климатическая система совершался за короткий отрезок времени (несколько тысяч лет). Крупнейший климатический скачок, произошедший на границе плейстоцена и голоцена, мог вызвать адаптивное напряжение в популяциях ключевых видов.
- В популяциях крупных фитофагов отмечены следы деградации, которые появлялись синхронно крупным климатическим скачкам, и особенно ярко они проявились на рубеже перехода плейстоцена к голоцену.
- Позднеплейстоценовые сообщества Восточноевропейской равнины представляли собой пастбищные экосистемы, объединявшие в единые ценозы виды, в настоящее время обитающие раздельно. Структурное единство пастбищных экосистем поддерживалось за счет средопреобразующей роли мегафауны и крупных копытных.
- Деструктивные проявления в популяциях крупных фитофагов ослабили её средопреобразующую роль и инициировали трансформацию позднеплейстоценовых пастбищных экосистем.
2.4. Рассчет антропогенной нагрузки на популяции пищевых доминантов для модельных ХКТ позднего палеолита Русской равнины
Хозяйственно - культурные типы были рассмотрены на примере модельных поселений. Для обеих моделей мною были произведены рассчеты антропогенной нагрузки на локальные популяции животных, представляющих собой главный пищевой ресурс: северного оленя и мамонта. При этом рассмотрены возможные сценарии воздействия палеолитических общин на популяции этих видов. При расчете антропогенной нагрузки на популяции пищевого доминанта для кочевой модели с ориентацией на стадных копытных были использованы современные данные о популяции северного оленя, как наиболее яркого представителя стадных копытных позднеплейстоценовых биоценозов, сохранившемся в естественных условиях почти до наших дней. (Егоров, 1965; Верещагин, Русаков, 1979; Сыроечковский, 1986,). При рассчете антропогенной нагрузки на популяции мамонтов, для оседлой модели были использованы эколого-популяционные характеристики для популяций африканских слонов (Бауэр, 1964; Слоны и другие гиганты.., 1983; Нири, 1984).
Кочевая модель.
Расчеты основаны на следующих допущениях:
1. Палеолитическая община модельного поселения не превышала 30 человек (Позднепалеолитическое..,1998).
2. Площадь освоения общины составляла 500 кмІ за год (Бромлей, 1986)
3. На указанной площади одновременно с общиной модельного поселения другие общины не проживали.
4. Северный олень составлял не менее 50% пищевого рациона общины (Позднепалеолитическое..,1974; Позднепалеолитическое поселение.., 1998)
5. Суточная энергетическая норма пищи на одного человека составляла не менее 3000 кал, т.е. на оленину приходилось 1500 кал. (Прохоров, 1994)
6. Изменения плотности оленей по сезонам повторяли современные схемы (лето 1 - 2 особей /кмІ, осенью и весной 5 особей /кмІ, зимой 6-7 особей /кмІ) (Сыроечковский, 1986; Верещагин, Русаков, 1974)
7. Изменение массы тела оленя по сезонам и калорийность оленины повторяла современные схемы (Сыроечковский, 1986; Таблицы химического состава и питательной ценности, 1961).
8. Убойный вес северного оленя был равен весу современных оленей, и составлял в среднем 70 килограмм (Сыроечковский, 1986)
9. Естественный прирост популяции позднеплейстоценового северного оленя был равен 14 -17% (в расчете использовалось среднее 15,5%), что характерно для современных популяций Центральной Якутии (Егоров 1965; Верещагин, Русаков,1974; Сыроечковский, 1986).
В таблице 2.4.1. представлены схема изменения физиологического состояния и калорийности мяса северного оленя по сезонам.
В зависимости от сезона происходило изменение суточной и месячной нормы в потреблении мяса.
Таблица 2.4.1.
Потребление мяса северного оленя на модельном поселении ХКТ кочевых охотников на стадных копытных
Сезон
(месяц)
|
Физиологическое состояние
северного оленя
|
Калорийность мяса
северного
оленя
(ккал/100г)
|
Суточная
норма
мяса на человека
(граммы)
|
Суточная норма
мяса на
общину
(кг.)
|
Месячное
потребление
мяса
общиной
(кг)
|
Соответствие
массы
потребленной
пищи числу
животных
(оленям)
|
август,
сентябрь, октябрь
|
упитанность
выше средней
|
276
|
558
|
16,7
|
502
|
6
|
ноябрь, декабрь, май, июнь, июль
|
средняя упитанность
|
138
|
1087
|
32,6
|
978
|
12
|
январь,
февраль,
март, апрель
|
упитанность
ниже средней
|
98
|
1510
|
45,3
|
1359
|
19
|
Мною рассмотрено 3 наиболее вероятных сценария, сезонного поведения палеолитической общины модельного поселения. Затем к каждому из них подобран современный этнографический аналог, описывающий долю используемого в пищу мяса от общего убойного веса северного оленя. Материалы для этнографических аналогий были заимствованы из работ Ю. Б. Симченко (1977), Е. Е. Сыроечковского (1986), Ф. Моуэт (1988).
1 сценарий: модельное поселении использовалось как зимнее стойбище.
Период временной оседлости не меньше 5 месяцев (октябрь, ноябрь, декабрь, январь, февраль, март). Диета на 100% состояла из мяса северного оленя.
1а сценарий: рассчитан при условии, что община использовала в пищу 75% теоретически возможного живого веса северного оленя.
1в сценарий: рассчитан при условии, что община использовала в пищу не более 50% теоретически возможного живого веса северного оленя
2 сценарий: модельное поселение использовалось как весенне-летнее стойбище. Период временной оседлости совпадает со временем массовых миграций северного оленя и составляет не более 3 месяца (апрель, май, июнь). Община использовала 100% мяса убитых северных оленей.
2а сценарий: рассчитан при условии, что община использовала в пищу 25% теоретически возможного живого веса северного оленя.
2в сценарий: рассчитан при условии, что община использовала в пищу 10% теоретически возможного живого веса северного оленя.
3 сценарий: модельное поселение использовалось как базовое летнее стойбище. Период временной оседлости совпадает с бесснежным периодом и составляет не менее 5 месяцев (апрель, май, июнь, июль, август, сентябрь). Община использовала 100% мяса убитых северных оленей.
3а сценарий: рассчитан при условии, что община использовала в пищу 50% теоретически возможного живого веса северного оленя.
3в сценарий: рассчитан при условии, что община использовала в пищу 25% теоретически возможного живого веса северного оленя
В таблице 2.4.2. , представлены возможные показатели оценки воздействия охоты на популяцию северного оленя.
Таблица 2.4.2.
Антропогенное изъятие северного оленя на модельном поселении ХКТ кочевых охотников на стадных копытных по разным сценариям
сценарии
|
Доля мяса использованного
в пищу (%)
|
Число
добытых
оленей (шт.)
|
Общее число
оленей за период (особей/500кмІ)
|
Процент изъятия
(%)
|
Прирост популяции северного оленя с учетом охоты (%)
|
1 сценарий
|
100
|
87
|
750
|
11,6
|
3,9
|
1а. сценарий
|
75
|
109
|
750
|
14,5
|
1
|
1в. сценарий
|
50
|
130
|
750
|
17,3
|
-2,2
|
2 сценарий
|
100
|
43
|
2500
|
2
|
13,5
|
2а. сценарий
|
25
|
75
|
2500
|
3
|
12,5
|
2в. сценарий
|
10
|
82
|
2500
|
3,3
|
12,2
|
3 сценарий
|
100
|
67
|
3000
|
2,23
|
13,27
|
3а. сценарий
|
50
|
101
|
3000
|
3,37
|
12,13
|
3в. сценарий
|
25
|
117
|
3000
|
4
|
11,5
|
Расчет антропогенного изъятия северного оленя на модельных поселениях ХКТ кочевых охотников на стадных копытных показал возможность палеолитических общин, специализирующихся в охоте на северного оленя нарушить устойчивость его популяции. Отрицательный прирост в популяции северного оленя (выделено), вызванный охотой на него мог наблюдаться на зимних стойбищах при использовании в пищу не более 50% мяса убитых животных от теоретически возможного (1в. сценарий). В расчетах не были учтены массовые случаи гибели северных оленей при природных катастрофах (болезни, голод, наводнение, и др.), что могло значительно сократить естественный прирост популяции, и как показывают, наблюдения Е. Е. Сыроечковского (1986), годовой прирост популяции в 2-4% часто эти потери не компенсирует. Таким образом, если даже принять, что на территории в 500 кмІ регулярно охотилась только одна община, специализированная охота на виды второго размерного класса на стойбищах зимнего периода, в некоторых случаях могла привести к снижению численности популяций главного объекта охоты. Материалы известные мне на настоящий момент, не позволяют говорить о возможности палеолитических общин с ХКТ кочевых охотников на стадных копытных, оказать заметное влияние на численность популяций промыслового вида в весенне-летний и летний период.
Стационарная (оседлая) модель.
Расчеты основаны на следующих допущениях:
1. Величина общины не превышала 40 человек (Первобытный человек, 1997).
2. Площадь освоения палеолитическими охотниками не превышала 500 кмІ
(Формозов, 1970)
3. Мясо мамонта составляло не менее 50% пищевого рациона общины
(Бромлей, 1986).
4. Суточная минимальная энергетическая норма для палеолитического человека составляла не менее 3000 ккал в сутки, тогда на мясо мамонта приходилось 1500 ккал/сут/человека (Будыко, 1977).
5. Убойный вес взрослого мамонта составлял не более 2000 кг, а неполовозрелого мамонта не превышал 1000 кг (Верещагин 1974; Бауэр, 1964).
6. Калорийность мяса мамонта составляла 200 ккал/100г. (Будыко, 1977, Первобытный человек, 1997).
7. Плотность популяции мамонтов для палеолита центра Русской равнины составляла не больше 1 особи /5 кмІ(Будыко, 1977).
8. Расчеты плодовитости проводились по данным для популяций современных африканских слонов (Слоны и другие гиганты…, 1982).
9. Ежегодный прирост популяции мамонтов при данных условиях составлял 7 особей/500 кмІ (Пидопличко (1951) оценил плодовитость мамонта в 0,08 %, что при указанной плотности популяции составляло годовой прирост в 8 особей/500 кмІ)
Количество животных, мясо которых восполняло суточные и годовые потребности общины в зависимости от возраста промыслового вида могло меняться. В таблице 2.4 3. представлена схема потребления мяса при условии, что в пищу используются неполовозрелые и взрослые особи. Разница в весе между ними составляет 50%.
Таблица 2.4.3.
Потребление мяса мамонта на модельном поселении ХКТ оседлых охотников на мамонта
Возрастное
состояние
|
Суточная норма
мяса на человека
в сутки
( кг.)
|
Суточная норма
мяса на общину
в сутки
(кг.)
|
Годовое
потребление мяса
общиной (кг.)
|
Соответствие
массы потребленной
пищи числу животных
(особей)
|
половозрелые
|
1,5
|
60
|
22000
|
11
|
неполовозрелые
|
1,5
|
60
|
22000
|
22
|
Мною рассмотрено два сценария описывающие варианты использования в пищу мяса мамонта, которые могли применяться при специализации в охоте, как на взрослых особей, так и на неполовозрелых.
1 сценарий: предполагает, что основу промысловой добычи составляли взрослые мамонты. Община использовала в пищу 100% убойного веса мамонта.
1а сценарий: рассчитан при условии, что община использовала в пищу 50 % мяса от убойного веса мамонта.
1в сценарий: рассчитан при условии, что община использовала в пищу 25 % мяса от убойного веса мамонта.
2 сценарий: предполагает, что промысловую добычу составляли неполовозрелые особи мамонта. Община использовала в пищу 100% убойного веса мамонта.
2а сценарий: рассчитан при условии, что община использовала в пищу 50 % мяса от убойного веса мамонта.
2в сценарий: рассчитан при условии, что община использовала в пищу 20 % мяса от убойного веса мамонта.
В таблице 2.4.4. представлены возможные оценки воздействия охоты на популяцию мамонта.
Таблица 2.4.4.
Антропогенное изъятие мамонта на модельном поселении ХКТ оседлых охотников на мамонта по разным сценариям
сценарий
|
Доля мяса, использованного
в пищу (%)
|
Число добытых
мамонтов (шт.)
|
Общее число
мамонтов за период (особей/500кмІ)
|
Процент изъятия
(%)
|
Прирост популяции мамонта с учетом охоты (%)
|
1 сценарий
|
100
|
11
|
100
|
11
|
-4
|
1а сценарий
|
50
|
17
|
100
|
17
|
-10
|
1в сценарий
|
25
|
19
|
100
|
19
|
-12
|
2 сценарий
|
100
|
22
|
100
|
22
|
-15
|
2а сценарий
|
50
|
33
|
100
|
33
|
-26
|
2в сценарий
|
25
|
38
|
100
|
38
|
-31
|
Расчет антропогенного изъятия мамонта на модельном поселении ХКТ оседлых охотников на мамонта показал, что доля животных потребляемая, человеком в результате охоты по всем сценариям превосходит естественный прирост популяции. При этом, для сценариев с ориентацией охоты на неполовозрелых особей (2, 2а, 2в сценарии) превышение естественного прироста столь значительно, что позволило бы общине за 3-4 года полностью истребить популяцию мамонтов в ареале своего обитания. Согласно сценариям с ориентацией в охоте на взрослых особей полное истребление популяции мамонтов в указанном ареале могло занять 10-25 лет. Следует вспомнить, что по данным археологов (об этом замечено выше) интервал существования поселений этого типа не превышал как раз 20 -25 лет. Вторая группа сценариев могла быть вполне вероятной. Анализ костного материала с ряда позднепалеолитических поселений Русской равнины показывает высокую долю неполовозрелых особей в “кухонных отходах”.
Таблица 2.4.5.
Возрастная структура популяций мамонтов с позднепалеолитических поселений и естественных захоронений
Местонахождение
|
Половозрелые
(до 1 гола)
|
Полувзрослые
(до10лет)
|
Взрослые
|
количество
|
доля (%)
|
количество
|
доля (%)
|
количество
|
доля (%)
|
Хотылево 2 (чертеж)
(Чубур, 1993)
|
7
|
4
|
63
|
36
|
105
|
60
|
Хотылево 2
Коллекция (Чубур, 1993)
|
1
|
2,2
|
16
|
34,8
|
31
|
63
|
Мезин (Чубур, 1993)
|
-
|
9
|
-
|
38
|
-
|
53
|
Берелех (Верещагин, 1972)
(природное захоронение)
|
-
|
-
|
-
|
32
|
-
|
_
|
Костенковско-Борщовский район (Верещагин, Кузмина, 1982)
|
-
|
5,6
|
-
|
16,5
|
-
|
76,9
|
Костенки 1 (Соффер, 1993)
|
-
|
-
|
-
|
68
|
-
|
32
|
Павлово 1 (Соффер, 1993)
|
-
|
-
|
-
|
60
|
-
|
40
|
Аналогии с африканским слоном, приведенные у Г. Бауэра (1965) позволяют заключить, что африканские аборигены так же часто выбирали для охоты неполовозрелых особей. Существование в позднем палеолите избирательной охоты могло нарушить возрастную и половую структуру популяций мамонта, что могло способствовать ускорению темпов вымирания мамонта.
Выводы
Основными хозяйственно-культурными типами Русской равнины являлись: ХКТ кочевых охотников на стадных копытных и ХКТ оседлых охотников на мамонта.
Рассчет антропогенного изъятия особей на модельных поселениях этих ХКТ, через определение минимальных потребностей суточной калорийности пищи, показал, что ХКТ оседлых охотников на мамонта существенно отличается от ХКТ кочующих охотников на стадных копытных.
Охота общины ХКТ кочующих охотников на стадных копытных специализировавшиеся на одном из видов могла способствовать снижению численности популяции пищевого доминанта, только при зимнем промысле и неполном использовании мяса убитых животных.
Охота общины ХКТ оседлых охотников на мамонта, могла способствовать снижению численности его популяции во все сезоны года при всех типах использования мяса убитых особей.
Охота на мамонта, с частичным использованием мяса животных в пищу, могла способствовать истреблению вида в границах ареала освоения этого ХКТ в короткие сроки (до 20 лет). Охота, ориентированная на неполовозрелых особей мамонта, была свойственна для ряда позднепалеолитических поселений Русской равнины, и могла привести к нарушению возрастной структуры популяций и ускорить вымирание мамонта на территории Русской равнины.
3.2. Изменения видового обилия костного материала основных видов “охотничьей фауны” на поселениях Восточно-Европейской равнины в течение пяти периодов позднего плейстоцена
В данной работе было рассмотрено изменение видового обилия основных видов охотничьей фауны (30 видов млекопитающих) на протяжении пяти периодов позднего плейстоцена. Для сравнения видового обилия костного материала использовался средний коэффициент обилия, который является отношением суммарного обилия вида со всех стоянок определенного периода к числу стоянок этого же периода.
Изменение обилия костного материала на поселениях Восточно-Европейской равнины в течение пяти периодов позднего плейстоцена представлено в приложении, рис. 3.2.1.
Для анализа были использованы виды эдификаторы и виды индикаторы климатических условий. Привожу краткое описание обилия костного материала этих видов.
Мамонт
Мамонт являлся главным объектом охоты на протяжении почти всего рассмотренного в работе периода. Обилие костного материала мамонта постепенно нарастает, достигнув максимума в интервале 18000 - 12000 лет назад. В настоящее время, нет оснований полагать, что численность и соответственно плотность популяции мамонта на Русской равнине в это время стабильно увеличивалась. Скорее можно предположить обратный сценарий, который по ряду палеогеографических реконструкций предполагает ухудшение условий обитания для этих животных (Динамика.., 2002). В таких условиях, на фоне естественного сокращения численности популяций, как ответ на изменение природной обстановки, активная охота человека могла активизировать вымирание мамонта на территории Восточно-Европейской равнины. Подтверждает это и резкое снижение обилия находок вида (в 6 раз) в последний из рассмотренных периодов. Столь катастрофическое снижение обилия мамонта можно объяснить совместным влиянием перепромысла человека и перестроек ландшафтов.
Северный олень
Северный олень входит в группу основных пищевых промысловых животных позднего плейстоцена. Обилие костного материала этого вида в течение рассмотренного времени стабильно увеличивается. Резкое увеличение обилия северного оленя - почти в три раза - в интервале 33000 - 24000 лет назад может быть объяснено переходом палеолитических охотников на специализированный промысел этого вида. Увеличение обилия северного оленя в рассмотренные периоды происходило одновременно с уменьшением обилия находок современных степных видов. Это может свидетельствовать о том, что заметную роль в изменении обилия и ареалов ряда видов играли и природные факторы. Максимальное увеличение обилия находок костного материала северного оленя отмечается 18000 - 12000 лет назад, вероятно, в самый неблагоприятный для существования позднеплейстоценовой фауны период вюрма, что может говорить о высокой пластичности и устойчивости этого вида. В этот период, снижается обилие находок всех рассмотренных в работе видов, кроме мамонта и северного оленя. По этой причине можно предположить для периода 18000 - 12000 лет назад наиболее неблагоприятные условия.
Лошадь
Лошадь - один из основных объектов охоты палеолитического человека. Обилие костного материала лошадей в рассмотренный период было подвержено изменениям. Максимальное увеличение находок наблюдалось в период 33000 -24000 лет назад, в последующий период - наиболее холодный - отмечается падение находок, продолжившееся резким снижением находок на границе плейстоцена и голоцена. Уменьшение обилия костного материала от лошадей во второй половине вюрма могло стать результатом преобразования пастбищных экосистем в лесные. Исследования экологии лошадей, сохранившихся до исторического времени (Боголюбский, 1959; Кириков, 1966), показывают, что оптимальными местообитаниями для вида являлись степи и лесостепи. Очевидно, в подобной природной обстановки позднего вюрма популяции лошадей на Русской равнине отличались высокой численностью, что и позволило этому виду стать одним из главных промысловых животных. Увеличение находок лошади 33000 - 24000 лет, на мой взгляд, следует связывать с увеличением промысла на нее.
Бизон
Бизон входил в группу основных объектов промысла палеолитического человека. Максимальное обилие находок этого вида наблюдается в южной половине Восточно-Европейской равнины. В настоящее время установлено, что оптимальные местообитания для бизона - степи (Боголюбский, 1959; Клаудели-Томпсон, 1982). В рассмотренный период обилие костного материала бизона стабильно снижается, при этом резкого падения обилия, как у многих других крупных фитофагов плейстоцена, не происходило. Очевидно, причиной снижения обилия находок костного материала бизона стало сокращение численности популяций этого вида, которое может быть объяснено теми же причинами, что и для лошади и ряда других копытных, предпочитающих открытые ландшафты.
Благородный олень
Один из древнейших промысловых видов на территории Восточно-Европейской равнины, однако, обилие находок большую часть рассмотренного времени оставалось низкоим. Объяснить это можно тем, что благородные олени редко образует крупные стада, а охота на одиночных особей не дает большого обилия костного материала. По этим причинам, рост обилия находок этого вида в течение позднего плейстоцена можно объяснить увеличением численности и плотности популяций благородного оленя. В холодные периоды вюрма обилие вида сокращается, в более теплые возрастает. Максимальное обилие находок отмечается 12000-9000 лет назад, когда лесистость равнины заметно возрастает, а комплекс позднеплейстоценовой фауны распадается.
Заяц
Обилие костного материала зайца на поселениях в позднем палеолите постепенно нарастает, при этом темпы увеличения наивысшее из числа рассмотренных в работе видов. Учитывая плохую сохранность костей мелких животных, можно предположить еще более высокую долю этого вида на поселениях. В повседневном рационе доля зайца, очевидно, была больше, чем об этом можно судить по палеонтологическим данным. Максимальное обилие вида достигается в интервале 12000 - 9000, и, может быть, связано не с увеличением численности популяций зайца, а с вынужденной сменой ориентации в охоте палеолитического человека.
Песец
Песец один из главных непищевых промысловых животных на территории Восточно-Европейской равнины. В рассмотренный период происходит увеличение обилия находок от среднего вюрма к интервалу 24000 - 18000 лет назад, что может быть связано с увеличением численности песцов на территории равнине, обусловленное похолоданием и развитием более благоприятной для него природной обстановки. В тоже время можно предположить, что численность песца за это время не менялась, а увеличение находок связано с интенсификацией охоты на этот вид, вызванной возросшими потребностями в мехе. Дальнейшее уменьшение обилия вида до голоцена, на фоне относительной стабильности такого же современного северного вида, как северный олень, может быть связано с перестройкой биоценотического покрова Русской равнины.
Пещерные хищники
В эту группу были включены пещерный лев, пещерная гиена, пещерный медведь. Обилие этих видов достигает значимых величин только в ранние периоды вюрма. После 33000 - 24000 лет назад обилие видов резко сократилось. На мой взгляд, для пещерной гиены и льва это связано со снижением численности мегафауны и крупных копытных, которое могло наблюдаться в этот неблагоприятный для них период. В изменившихся природных условиях более разнообразная специализация, как у волка, оказалась выгоднее, и позволила ему доминировать в составе непищевой охотничьей фауне.
3.3. Анализ изменения ареалов и обилия костного материала основных экологических группировок “охотничьей фауны” поселений Восточно-Европейской равнины
В исследованиях позднепалеолитического фаунистического комплекса принято делить его на 4 экологические группировки: тундровые, степные, лесные, вымершие виды (Дашкевич, 1969; Верещагин, 1974; Хотинский, 1981). На этой основе виды “охотничьей фауны”, информация о которых была собрана в БД, были разделены на 4 аналогичные группы, представленные в таблице 3.3.1.
Таблица 3.3.1.
Экологические группировки видов “охотничьей фауны”
на позднепалеолитических поселениях Восточно-Европейской равнины
Экологическая группировка
|
Виды или роды
|
Число таксонов
|
тундровые
|
Rangifer tarandus Smith - (северный олень)
Ovibos moschatus Zimm - (мускусный овцебык)
Alopex lagopus L - (песец)
Disrostonx Gloger - (копытные лемминги)
Lemmus Link - (лемминги обыкновенные)
|
5
|
лесные
|
Bos primigenius Boj - (первобытный тур)
Capreolus capreolus L - (косуля обыкновенная)
Alces alces L - (лось)
Cervus elaphus L - (благородный олень)
Canis L - (волк), Gulo gulo L - (росомаха)
Ursus arctos L - (медведь бурый), Lepus L - (заяц)
Vulpes vulpes L - (лисица обыкновенная)
Martes martes L - (куница лесная)
Meles meles L - (барсук обыкновенный)
Castor fiber L - (бобр речной)
Mustela nivalis L - (ласка)
Mustela zibellina L - (соболь)
Mustela erminea L - (горностай)
Mustela putorius L - (хорек лесной)
Desmana Guld - (выхухоль)
Arvicola terrestris L - (водяная крыса)
Lutra lutra L - (выдра речная)
Felis lynx L - (рысь), Sus scrofa L - (кабан)
|
21
|
степные
|
Bison priscus Boj - (бизон первобытный)
Equus L - (лошадь), Saiga tatarica L - (сайгак )
Vulpes corsak L - (лиса-корсак)
Ochotona Link - (пищуха), Marmota Blum - (сурок)
Alloctaga F Cuvier - (тушканчик), Citellus Oken - (суслик)
Cricetus Milne-Edwards - (хомяк)
Allocricetulus Argyr - (хомячок)
Ellobius Waldh - (слепушонка), Spalax Guld - (слепыш)
Equus Hydruntinus Regali - (плейстоценовый осел)
|
13
|
вымершие
|
Mamuthus primigenius Blum - (мамонт поздний)
Coleodonta antiquitatis Blum - (волосатый носорог)
Megaloceros giganteus Blum - (большерогий олень)
Ursus spelaeus Rosenm - (пещерный медведь)
Hyaena spelaea - (пещерная гиена)
Panthera spelaea Gold - (пещерный лев)
|
6
|
Необходимо, отметить, что подобное деление на экологические группировки для вводов позднеплейстоценовой фауны носит, условный характер. Для вымерших видов, экологические особенности до настоящего времени достоверно не установлены. В тоже время, по мнению многих исследователей (Пидопличко, 1954, Верещагин, 1979, Калякин, Турубанова, 2002) подобное разделение на группы плейстоценовой фауны, может дать общее представление об особенностях ландшафтов времени обитания вида.
Тундровая группировка
Виды, составляющие в настоящее время тундровую экологическую группировку, в рассмотренный период имели более широкий ареал, чем в современное время.
Распространение и обилие тундровых видов в “охотничьей фауне” поселений Восточно-Европейской равнины представлено в приложении, рис. 3.3.1.
До интервала 18000 - 12000 лет назад ареал некоторых тундровых видов доходил до побережья Черного и Азовского моря. При этом высота горных стоянок Крыма и Кавказа, где были отмечены костные остатки тундровых видов, чаще всего не превышает 600 - 800 метров, что не позволяет сделать предположение о существовании этих видов в нивальной зоне. Рассматривая этот вопрос, некоторые исследователи (Четвертичный период 1965) указывают на возможность дальних заходов на юг северных оленей и песцов, однако, остатки тундровых грызунов были найдены так же далеко к югу от их современного ареала (окрестности Брянска, Новгорода - Северского). Ареал тундровых видов не изменился в относительно теплый брянский интервал (33000-24000 л.н.), а в более холодный “поздневюрмский пессимум ” (24000 18000) даже немного сократился. Резкое сокращение ареала видов произошло уже в голоцене (12000 - 9000), после окончательной трансформации смешанных вюрмских биоценозов. Увеличение обилия тундровых видов, судя по костному материалу, произошло в интервалы 33000 - 24000 л.н. и 24000 - 18000 л. н., что можно связать с общим похолоданием на территории Восточно-Европейской равнины. В это время ранее равномерно распределенные находки, оказались локализованы в центре равнины. Изменение обилия отдельных тундровых видов, за исключение овцебыка, повторяет общую схему для видов тундровой группировки в целом.
Обилие находок песца в “охотничьей фауне” позднего палеолита Русской равнины представлено в приложении, рис. 3.3.2.
Лесная группировка
Виды, составляющие в настоящее время лесную группировку, в течение всего рассмотренного периода имели обширный ареал, охватывавший всю территорию Восточно-Европейской равнины, за исключением крайнего северо-запада (приложение, рис. 3.3.3).
Материалы, собранные в базе данных не позволяю выделить заметных сдвигов ареалов лесных видов в рассмотренные периоды. Подобная стабильность ареалов животных данной экологической группы, может быть объяснена амфибореальным распространением лесных массивов в вюрме (Палеогеография.., 1978). В отсутствии единой лесной зоны, вюрмские климатические пульсации не нашли заметного отражения в изменении ареалов животных лесного комплекса. Обилие находок лесных видов в рассмотренное время нарастает, что может говорить о постепенном увеличение лесистости Восточно-Европейской равнины от раннего и среднего вюрма к голоцену. Стабильно высока концентрация местонахождений, с обилием костного материала лесных видов в течение всего позднего вюрма, оставалась в предгорьях Кавказа, Крыма, южного Урала, Молдавии. В этих районах могли существовать относительно крупные лесные массивы вюрма. Изменение обилия отдельных лесных видов в рассмотренный период в целом можно описать единой схемой. Стабильное увеличение количества находок во все рассмотренные периоды характерно для лося, бурого медведя, зайца, лисицы и др.
Обилие находок лося в “охотничьей фауне” позднего палеолита Русской равнины представлено в приложении, рис. 3.3.4.
Для большинства из них рост обилия происходил с нарастающей динамикой. Обилие лося, лисицы, кабана и косули в последний рассмотренный период увеличилось в 2 - 4 раза, по сравнению с предшествующим. Подобные особенности изменения обилия отдельных видов могут свидетельствовать о высоких темпах роста численности популяций и освоения новых территорий лесными видами после распада мамонтового фаунистического комплекса.
Степная группировка
Изменения ареалов видов, слагавших в историческое время степную группировку, в позднем плейстоцене не было однозначным.
Распространение и обилие степных видов в “охотничьей фауне” поселений Восточно-Европейской равнины представлено в приложении, рис. 3.3.5.
Ареал сокращался, смещаясь к югу с раннего и среднего вюрма (поселения старше 33000 л. н.), когда находки степных видов известны для территорий современных республики Коми, Среднего Урала, центральной Беларуси. В два последующих периода северная граница стабилизируется. В заключительные этапы плейстоцена, и начале голоцена степные виды снова продвигаются к северу. Объяснить подобные изменения ареала видов этой группы можно похолоданием в позднем плейстоцене, что могло сместить ареалы некоторых видов к югу. Однако реакция степных видов на эту климатическую пульсацию отчетливо не прослеживается. Большую часть рассмотренного времени природная обстановка на территории равнины благоприятствовала видам предпочитающим слабо облесенные ландшафты, а климатические пульсации, видимо, не являлись для них лимитирующим природным фактором. Пространственное размещение находок степных видов показывает сдвиг обилия и количества местонахождений степных видов к югу, что может быт следствием большей численности и плотности популяций этих видов в этих районах. Изменение обилия отдельных представителей современной степной группировки более ярко указывает на уменьшение обилия находок степных видов от раннего вюрма к началу голоцена. Особенно четко это прослеживается для сайги, бизона, плейстоценового осла.
Обилие находок сайги в “охотничьей фауне” позднего палеолита Русской равнины представлено в приложении, рис. 3.3.6.
Для степных грызунов ситуация менее однозначная или вовсе противоположная, хотя количество находок этих видов не достаточно для объективного анализа.
Группировка вымерших животных
Оптимальные экологические условия дл этой группы животных достоверно еще не установлены. Некоторые исследователи считают, что многие из них были космополитами (Верещагин, 1979). Данные, собранные в БД так же указывают на широкий спектр природных условий, в которых могли обитать вымершие виды.
Распространение и обилие вымерших видов в “охотничьей фауне” поселений Восточно-Европейской равнины представлено в приложении, рис. 3.3.7.
Ареал этих видов во все периоды вюрма, за исключением последнего к которому многие из этих видов вымерли, был обширным. Сдвигов его северных и южных границ не установлено, однако к последнему интервалу (12000 - 9000) единый ареал вымерших видов распался на мелкие локусы, в которых произошло окончательное вымирание этих видов. Обилие костного материала эти животных (прежде всего мамонта) на поселениях зависело, главным образом, не от численности и плотности видов в окрестностях, а от успехов охоты или собирательства костей.
Обилие находок пещерный медведь в “охотничьей фауне” позднего палеолита Русской равнины представлено в приложении, рис. 3.3.8.
В изменении обилия отдельных вымерших видов (пещерных хищников), прослеживаются черты направленного вымирания, так как их обилие в находках стабильно сокращается до нуля. Сокращение обилия крупных пещерных хищников могло быть инициировано схожими процессами в популяциях мегафауны.
На рис. 3.3.9. (приложение) представлен график изменения обилия костного материала рассмотренных групп.
На поселениях старше 33000 лет назад отмечается максимальное обилие степных видов и вымерших видов с неясной экологией, в тоже время обилие лесных и тундровых видов минимально. Подобное соотношение экологических групп животных, может свидетельствовать об относительно теплом и сухом климате на территории Восточно-Европейской равнины в интервале 70000 - 33000 лет назад. Преобладающим типом биоценозов очевидно, являлись степи и лесостепи. В брянское время (24000-18000 лет назад) сокращается обилие степных видов и вымерших видов не ясной экологии, а обилие лесных и тундровых возрастает. Очевидно, с этого времени, начинается преобразование вюрмских ценозов, которое могло быть инициировано резким климатическим скачком, изменившим климатические условия в сторону похолодания и увеличения увлажнения. Для времени поздневюрмского пессимума, изменения соотношения экологических групп животных в охотничьей фауне подтверждают мнения о том, что климатические условия этого времени были максимально холодными и ксерофитными. Доля степных видов в охотничьей фауне к этому времени, сократившаяся более чем в два раза, стабилизируется. Доля лесных видов стабильно увеличивается до начала голоцена. К этому времени относится максимальная доля современных тундровых видов в охотничьей фауне. Доля вымерших видов не ясной экологии к этому времени стабилизируется, хотя тренд к снижению обилия этих видов наметился еще в среднем вюрме. Подобную приостановку в падении обилия этих видов, по-видимому, можно объяснить увеличением охоты на них (прежде всего на мамонта), что могло исказить состав охотничьей фауны, который к этому времени, вероятно, уже не отражал естественное состояние популяций. Таким образом, увеличение масштабов охоты на мамонта происходило на фоне естественного сокращения численности этого вида, вызванное изменением природной обстановки. Преобразование состава экологических групп животных охотничьей фауны в послеледниковье (12000 - 9000 лет назад) указывает на резкие изменения состава зооценозов. Увеличение обилия лесных видов происходит в противофазе с сокращением обилия вымерших видов с неясной экологией. На мой взгляд, между этими процессами есть взаимосвязь. Сокращение количества эдификаторов пастбищных экосистем ускорило их трансформацию в лесные сообщества.
3.4. Анализ изменения соотношений животных основных трофических уровней “охотничьей фауны” Восточно-Европейской равнины
Виды, информация по которым собрана в БД, разделены на 4 основные группы по особенностям питания и поведения.
Таблица 3.4.1.
Основные Группы животных “охотничьей фауны” Восточно-Европейской равнины по особенностям питания и поведения
Экологическая группа
|
мегафауна
|
Хищники (крупные)
|
Стадные копытные
|
Не стадные копытные
|
виды
|
Mamuthus primigenius - (мамонт поздний)
Coleodonta antiquitatis - (волосатый носорог)
|
Hyaena spelaea - (пещерная гиена)
Panthera spelaea - (пещерный лев) Canis (волк)
Gulo gulo - (росомаха)
Felis lynx - (рысь)
Alopex lagopus - (песец)
|
Bison priscusj - (бизон первобытный)
Equus - (лошадь)
Saiga tatarica - (сайгак )
Equus hydruntinus - (плейстоценовый осел)
Rangifer tarandus - (северный олень)
|
Megaloceros giganteus - (большерогий олень)
Capreolus capreolus - (косуля)
Alces alces - (лось)
Cervus elaphus - (благородный олень)
Sus scrofa - (кабан)
|
Для сравнения экологической структуры охотничьей фауны Восточно-Европейской равнины были использованы аналогичные данные Р. Фоули (1990) для Восточной Африки. В работе Р. Фоули, подсчет соотношений экологических групп производился при пересчете по видам. В таблице, приведенной ниже, дана доля видов каждой экологической группы от общего числа видов. Для плейстоценовой фауны Восточно-Европейской равнины палеонтологические материалы часто не позволяют произвести определение до вида, поэтому соотношения экологических групп для них были подсчитаны при пересчете их обилий. В таблице дана доля суммарного обилия таксонов каждой экологической группы от общего обилия всех таксонов.
Таблица 3.4.2.
Сравнение состава экологических групп животных охотничьей фауны позднепалеолитических поселений Восточно-Европейской равнины с современной и ископаемой фауны Восточной Африки (%)
Экологическая группа
|
Восточно-Европейская равнина
|
Восточная Африка
|
старше 33000 лет назад
|
33000 - 24000 лет назад
|
24000 - 18000 лет назад
|
18000 - 12000 лет назад
|
12000 - 9000 лет назад
|
Ископаемая (плейстоцен) (Фоули, 1990)
|
Современная
(Фоули, 1990)
|
мегафауна
|
15,4
|
13,4
|
20,8
|
15,6
|
5,2
|
25
|
11,9
|
хищники
|
13,3
|
18,6
|
18,3
|
13,9
|
13,8
|
12
|
16,6
|
стадные копытные
|
34,8
|
30,4
|
30,4
|
21,7
|
24,3
|
35,2
|
49,3
|
не стадные копытные
|
6,8
|
8,2
|
7,5
|
6,7
|
23,5
|
-
|
-
|
Сравнение экологической структуры охотничьей фауны Восточно-Европейской равнины с современной и ископаемой Восточной Африки, показывает их общее структурное сходство. Большую часть рассмотренного времени, экологическая структура охотничьей фауны Восточно-Европейской равнины, оставалась количественно близкой с современной и ископаемой для Восточной Африки. Экосистемы Восточно-Европейской равнины, почти в течение всего вюрма оставались пастбищными, что подтверждает, высока доля мегафауны и стадных копытных в них. Доля мегафауны в них была даже выше современной для Восточной Африки. В современной Восточной Африке господствующим типом растительных сообществ является саванны, которые многие исследователи считают типичными пастбищными экосистемами. Следует отметить, что структура палеосообществ Восточно-Европейской равнины существенно не менялась при многократных и часто “катастрофических”, как считают некоторые исследователи (об этом в главе 1) климатических пульсациях.
Структура экосистем Восточной Европы не менялась, в течение всего вюрма, в противном случае это нашло бы отражение в составе фауны. Повышение доли мегафауны в интервале 24000 - 18000 лет назад, объясняется резким увеличением охоты на неё, по этой причине структура охотничьей фауны оказалась искажена, по сравнению с экологической структурой естественных зооценозов. Самые крупные преобразования экологической структуры произошли в начале голоцена (12000 - 9000 лет назад) и затронули они, главным образом, мегафауну, доля которой сократилась в 3 раза. Освободившиеся пищевые ресурсы освоили не стадные копытные, доля которых увеличилась почти в 4 раза. Не стадные копытные это преимущественно лесные, оседлые виды и рост их доли в экологической структуре фауны может свидетельствовать о серьезных преобразованиях экосистем.
3.5. Пространственный анализ изменения состава доминантов в охотничьей фауне поселений Восточно-Европейской равнины в разные периоды вюрма
Определение доминантов в составе охотничьей фауны проводилось по обилию костного материала. Выявление преобладающих промысловых видов, по мнению А. А. Величко и И. Е. Куренной (1994), позволяет предположить специализацию в охоте палеолитических общин. Состав доминантов в разные периоды вюрма, для различных районов равнины не был однородным. Из 52 видов охотничьей фауны собранной в БД в разных районах Восточно-Европейской равнины по обилию костного материала преобладали 6 видов: мамонт, лошадь, северный олень, бизон, пещерный медведь и благородный олень. На территории Восточно-Европейской равнины, по особенностям локализации поселений, автором было выделено 9 районов. В течение рассмотренного в работе периода, число районов не было постоянно. Максимальное число районов (9) отмечалось 18000 - 12000 лет назад, для других временных интервалов, собранный материал, не позволил провести полное районирование. Для каждого из них были вычислены доли преобладающих видов по обилию костного материала в составе охотничьей фауны. Доля каждого вида определялась как отношение обилия вида на поселениях в приделах установленного района к общему обилию всех видов охотничьей фауны, обнаруженных на поселениях этого района.
Предкавказье (34 поселения)
Доля основных промысловых видов в охотничьей фауне Предкавказья представлено в приложении, рис. 3.5.1.
Для поселений старше 33000 лет назад характерна смешанная специализация в охоте. Примерно одинаковые доли в составе охотничьей фауны имеют 4 вида: лошадь, мамонт, бизон, пещерный медведь. В следующий период 33000- 24000 лет назад, в составе охотничьей фауны преобладаю лошадь и пещерный медведь. На эти виды приходится по ј от суммарного обилия всех видов, что может свидетельствовать о специализации в охоте на лошадь и пещерного медведя. На поселениях возрастом 24000 - 18000 лет назад монодоминантом охотничьей фауны становится пещерный медведь, который по обилию костного материала в 2 раза превосходит идущего на втором месте благородного оленя. На поселениях 18000 - 12000 лет назад ярко выраженная специализация отсутствует. По обилию костного материала в охотничьей фауне преобладают бизон (1/4) и благородный олень (1/5). Доля пещерного медведя незначительна. Подобное резкое снижение обилия костного материала пещерного медведя, может быть, связана с сокращением численности этого вида вследствие перепромысла.
Бассейн Десны (26 поселений)
Доля основных промысловых видов в охотничьей бассейна Десны представлено в приложении, рис. 3.5.2.
Для поселений этого района старше 33000 лет назад характерна смешанная специализация в охоте, с преобладанием по обилию бизона (27% обилия всех видов), мамонта (20%) и лошади (17%). Для периода 24000 - 18000 лет назад в составе охотничьей фауны преобладали мамонт (26%), северный олень (19%) и лошадь (16%). Таким образом, специализация в охоте сталась смешанной, не смотря на изменения состава доминантов. Мамонтовая специализация в районе могла существовать в период 18000 - 12000 лет назад, когда обилие костного материала мамонта достигает 33% от общего обилия всех видов.
Крым (12 поселений)
Доля основных промысловых видов в охотничьей фауне Крыма представлено в приложении, рис. 3.5.3.
Для поселений старше 33000 лет назад характерна специализация в охоте на лошадь, доля которой составляет 44% от обилия всех видов на поселениях этого района старше 33000 лет назад. Для группы поселения возрастом 24000 - 18000 и 18000 - 12000 лет назад специализация выражена менее ярко. На долю лошади приходится 28 процентов от общего обилия видов.
Бассейн Днестра (44 поселения)
Доля основных промысловых видов в охотничьей фауне бассейна Днестра представлено в приложении, рис. 3.5.4.
Для поселений старше 33000 лет назад в составе охотничьей фауны преобладают мамонт, лошадь, бизон, северный олень, доля этих видов примерно равна, что говорит о смешанной специализации в охоте для населения поселений этого периода. Для поселений возрастом 33000 - 24000 лет назад по обилию костного материала преобладают северный олень и лошадь, однако, доминирование этих видов не столь очевидно, чтобы делать предположения о специализации охоты на эти виды. Для поселений возрастом 24000 - 18000 лет назад специализация осталась смешанной с преобладанием мамонта (24%) по обилию костного материала. Для периода 18000 - 12000 лет назад характерно преобладание северного оленя в охотничьей фауне.
Средний Дон (62 поселения)
Доля основных промысловых видов в охотничьей фауне Среднего Дона представлено в приложении, рис. 3.5.5.
Для поселений этого района в интервале от 33000 до 12000 лет назад характерно преобладание в охотничьей фауне лошади и мамонта. Максимальная доля, которых отмечается для лошади (29,1%) в период 33000 - 24000 лет назад, а для мамонта (29,7%) 24000 - 18000 лет назад.
Предуралье и Северо-Восток Русской равнины (14 поселений)
Доля основных промысловых видов в охотничьей фауне Северо-востока Русской равнины представлено в приложении, рис. 3.5.6.
Для поселений возрастом 33000- 24000 лет назад характерно преобладание в охотничьей фауне мамонта и северного оленя. Доля лошади так же высока и лишь немного уступает двум первым видам. Для периода 18000 - 12000 лет назад более ярко проявилась специализация в охоте на мамонта и северного оленя.
Бассейн Днепра (16 поселений)
Мамонтовая специализация в охоте палеолитических общин этой области проявилась в интервале 24000 - 18000 лет назад. Для поселений возрастом 18000 - 12000 лет назад мамонтовая специализация проявилась максимально ярко. Доля мамонта в обилии костного материала всех видов достигает 45%.
Приазовье (5 поселений)
Для этой области в интервале 24000 - 12000 лет назад характерна специализация в охоте на бизона, доля которого составляет 37% от обилия всех костей, что всего лишь на 9% меньше чем обилие костного материала от лошади.
Северо-запад современной Украины (7 поселений)
Для поселений этого района в интервале 24000 - 12000 лет назад, характерна максимально высокая доля мамонта в охотничьей фауне среди всех районов Восточно-Европейской равнины, которая составляет 52%.
Таблица 3.5.6.
Особенности специализации в охоте для позднепалеолитических поселений разных районов Восточно-Европейской равнины
Район
|
Тип специализации охоты на поселениях разного возраста (лет назад)
|
Старше 33000 33000-24000 24000-18000 18000-12000
|
Предкавказье
|
Смешенная
|
Смешенная
|
Пещерный медведь
|
Смешенная
|
Бассейн Десны
|
Смешенная
|
-
|
Мамонт, не ярко
|
Мамонт
|
Крым
|
Лошадь
|
-
|
Лошадь
|
Бассейн Днестра
|
Смешенная
|
Смешенная
|
Смешенная
|
Северный олень и лошадь, не ярко
|
Средний Дон
|
-
|
Лошадь и мамонт, не ярко
|
Лошадь и мамонт
|
Лошадь и мамонт
|
Северо-восток Русской равнины
|
-
|
Смешенная
|
Смешенная
|
Бассейн Днепра
|
-
|
-
|
Мамонт, не ярко
|
Мамонт
|
Приазовье
|
-
|
-
|
Бизон, не ярко
|
Северо-запад Украины
|
-
|
-
|
Мамонт
|
-
|
На территории Восточно-Европейской равнины в разные периоды вюрма выделяется до 7 районов со своеобразным составом доминантов охотничьей фауны, что может свидетельствовать о разных типах охотничьей специализации существовавших на территории равнины в течении вюрма. Следует отметить, что охотничья специализация для большинства районов проявилась не ярко. Только для 4 районов в течении вюрма, доля пищевого доминанта превышает 40%: Крым (поселения старше 33 т. л. н., лошадь), Предкавказье (24 - 18 т. л. н., пещерный медведь), северо-восток современной Украины (24-18 т. л. н., мамонт) и бассейн Днепра (18-12 т.л.н., мамонт). Признаки специализированной охоты существуют уже на поселения возрастом старше 33000 лет назад. Наибольшее распространение специализированная охота получила в период 24000-18000 лет назад, когда выделяются области с преобладанием в охотничьей фауне: пещерного медведя, мамонта, лошади и бизона. На всех стоянках районов, где в охотничьей фауне по обилию костного материала доминирует мамонт, его содоминантом являлся северный олень. Подобная особенность может свидетельствовать о том, что специализация на мамонта возрастала в холодные интервалы вюрма, когда условия способствовали увеличению обилия современного северного вида. Наиболее типичная ситуация на позднепалеолитических поселениях Восточно-Европейской равнины, это преобладание в охотничьей фауне сразу 3-4 видов животных (мамонт и стадные копытные).
Выводы
Состав основных промысловых видов на протяжении рассмотренного времени оставался стабилен. Максимальное обилие костного материала в охотничьей фауне поселений Восточно-Европейской равнины приходится на мегафауну (мамонт и волосатый носорог) и стадных копытных.
Моновидовая специализация в охоте для большинства районов Восточно-Европейской равнины была слабо выражена. Чаще всего в составе охотничьей фауны одновременно преобладают несколько видов животных, доля которых не превышает 20-25% от обилия костного материала всех видов.
Экологический состав видов, слагавших охотничью фауну на протяжении всего рассмотренного времени, был смешанным. Однако обилие костного материала видов основных экологических групп в разных районах не было одинаково и также изменялось во времени.
В течение позднего плейстоцена происходило постепенное сокращение доли современных степных видов и вымерших видов неясной экологии. В тоже время, доля современных лесных видов постепенно возрастала и резко увеличилась на рубеже голоцена и плейстоцена.
Состав основных трофических и этологических групп млекопитающих позднего плейстоцена Восточно-Европейской равнины имел принципиальные сходства с аналогичным составом современной и ископаемой фауны Восточной Африки.
Пространственные изменения ареалов и количественные перераспределения видов охотничьей фауны, инициированные глобальными климатическими перестройками, начались уже в среднем вюрме и протекали постепенно на всем протяжении позднего плейстоцена. Усиление охотничьего промысла со стороны человека во второй половине вюрма активизировало перестройку позднеплейстоценовых ценозов и способствовало вымиранию наименее плодовитых из них.
Сокращение и последующее полное вымирание мегафауны способствовало потере позднеплейстоценовыми экосистемами своих структурных особенностей. Деградация пастбищных экосистем протекала на фоне совместного действия природных и антропогенных факторов, что существенное ускорило и усилило негативный эффект этих изменений на биоту Восточно-Европейской равнины.
Приложение
Рис. 1.1. Вюрмские местонахождения охотничьей фауны на территории
Восточной Европы.
Возраст местонахождений.
старше 32000 лет назад
32000-24000 лет назад
24000-18000 лет назад
18000-14000 лет назад
14000-10000 лет назад
Природные зоны времени “поздневалдайского
пессимум” (22000 - 18000 лет назад).
возраст
тысяч
лет
назад
|
Флон,, 1980
|
Маркова, 2002
|
Зубаков, 1986
|
Динамика
почвенных,, 1999
|
Спиридонова,
1991
|
Природа и
Человек, 1984
|
Лазуков, 1989
|
Свитич, 1987
|
Андреичева,
2002
|
Динамика..,
2002
|
Бердников,
1983
|
8
|
|
|
|
|
|
|
|
|
|
|
|
12
|
16
|
20
|
24
|
28
|
32
|
36
|
40
|
44
|
48
|
52
|
56
|
60
|
64
|
68
|
Рис.1.2. Термические ритмы некоторых интервалов второй половины вюрма
относительное потепление
относительное похолодание
нет данных
.
Рис. 1.5.1. Эколого-ценотический анализ состава перигляциальной флоры
А. Эколого-ценотический анализ ископаемой Б. Эколого-ценотическая перигляциальной флоры по Новенко, 2000. приуроченность компонентов ископаемой (22 - 18 тыс. лет назад) флоры бассейна Ср. Днестра
(15 -13 тыс. лет назад) (Среда обитания…, 1999)
Рис. 1.5.2.Распределение радиоуглеродных датировок остеологических материалов по мамонту для северной Евразии
(по данным: Исследование по современным и плейстоценовым млекопитающим (1995)).
Рис. 1.5.3. Изменение высоты тела половозрелых особей Mammuthus primigenius в позднем палеолите. (составлено по материалам А.О. Аверьянов (1985), В.В. Украинцева (1991), И.Е. Кузьмина, Н.Д. Праслов (1992), Н.А. Тихонов (1995))